![]() Pas de frame de navigation? Cliquez ici!
Pas de frame de navigation? Cliquez ici!
|
|
Il règne encore beaucoup de confusion autour de la notion de consanguinité. C'est pourtant une notion fondamentale pour tout élevage animal (mais aussi pour la sélection végétale). Ces notions font appel aux bases de la génétique et à une certaine formation scientifique (niveau Terminale S à DEUG de bio). Avant d'aller plus loin, il vous sera donc nécessaire de maîtriser à peu près les notions évoquées dans les parties "génétique" de Ratlala. Cette partie vous familiarisera avec l'approche moléculaire de la génétique : les gènes et leur expression.
Pour tout complément d'information, n'hésitez pas à consulter le forum Café Raoul.
Les différents points abordés :
![]()
|
|
Pour aborder la notion de consanguinité, nous allons fréquemment changer d'échelle. On ne s'intéressera plus uniquement à chaque gène, ni à chaque chromosome, ni même à l'individu. Ce qui nous intéressera surtout, c'est la génétique des populations, c'est à dire la structure génétique de chaque individu plus la structure génétique de l'ensemble des individus. C'est cela qui va influer sur la consanguinité et les relations génétiques entre les membres d'une population : échanges de gènes (reproduction), gènes en commun (liens parentaux), nouvelles mutations, divergences génétiques, etc.
On distingue deux notions légèrement différentes : deux individus sont apparentés si ils ont au moins un ancêtre en commun (lien entre deux individus) ; un individu est dit consanguin si ses deux parents sont apparentés (propre à un seul individu). Le niveau de consanguinité découle donc du degré de parenté. Si on remonte suffisamment loin, cela implique que tous les individus d'une espèce donnée sont consanguins, bien sûr. Cependant, on considère qu'au delà d'un certain seuil de "dilution" il n'y a plus de consanguinité significative. Statistiquement, plus deux rats sont apparentés, plus ils auront de gènes en commun, et plus leurs enfants seront consanguins. On peut relier plus ou moins directement le nombre de gènes en commun et le degré de parenté.
On parle de consanguinité pour un seul gène, mais aussi pour un
génotype complet. Ainsi, le fils hérite de 50% du patrimoine génétique de
sa mère et 50% de son père. Il a donc une chance sur deux d'hériter d'un
gène donné de son père (un seul gène) et il hérite de la moitié des
gènes de son père (génotype). Il est à 50% consanguin avec son père et à
50% consanguin avec sa mère. A chaque génération le taux de consanguinité
est donc divisé par deux, puisque chaque parent transmet la moitié de son
patrimoine génétique. Le petit-fils hérite donc (statistiquement) de 25% du
patrimoine génétique de chacun de ses grands-parents, 12,5% de celui de
chacun de ses arrières grands parents, etc.
C'est pourquoi chez les chevaux on parle de demi-sang, de quart de sang
"arabe", par exemple.
|
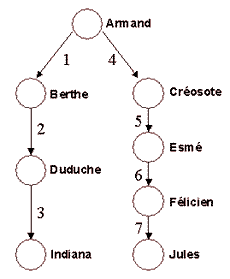
Le coefficient de parenté entre Indiana et Jules
est de: |
Le coefficient de consanguinité (Cc) : méthode de calcul Le coefficient de parenté est la probabilité pour que deux gènes
homologues tirés au hasard chez les individus I et J soient identiques.
Cc est la probabilité pour que deux gènes
homologues soient identiques chez l'individu K, leur fils. Considérons
que I et J aient un ancêtre commun, l'individu A. Ils sont donc
apparentés. A a transmis la moitié de ses gènes à chacun de ses
enfants, B et C. Eux aussi transmettent la moitié de leurs gènes à
leurs descendants. A chaque génération, la probabilité de retrouver
ce gène diminue donc de moitié. Attention, cette probabilité assure que les deux gènes viennent de A, pas que ce sont les mêmes, car les gènes de A vont par paires... Il faut donc encore diviser par deux pour garantir l'identité des deux gènes, si on considère que A n'est lui même pas consanguin. Sinon la probabilité devient 1/2 + Cc(A)/2 Quelques coefficients de parenté (et donc les taux de consanguinité de leurs enfants):
|
||||||
Les régimes de reproduction s'appliquent généralement à un seul gène ou à un groupe de gènes. On peut extrapoler plus ou moins facilement pour en déduire la structure du génome dans son ensemble, c'est à dire pour appliquer leurs conséquences aux individus des générations suivantes, mais on verra plus loin que cela présente quelques difficultés... Attention, ces régimes de reproductions sont des modèles théoriques. Ils ne s'appliquent que dans le cadre de populations illimitées (ou presque, disons un million d'individus ou plus, quoi...)
L'autogamie :
Ce régime de reproduction n'existe guère que chez les plantes et
quelques animaux parthénogénétiques. Dans ce cas de figure chaque individu
se féconde lui-même. Quand il se produit une mutation où A devient a, l'animal mutant, avec
un génotype Aa, donne 1/4 de AA, 1/4 de aa, et 1/2 de Aa. A chaque
génération il y a deux fois moins d'individus hétérozygotes. Très
rapidement les animaux issus de ce régime de reproduction deviennent
entièrement homozygotes, sauf lorsqu'une nouvelle mutation apparaît (ce qui
est plutôt rare). Lorsque la population est constituée d'individus
homozygotes pour tous leurs gènes, on obtient une lignée pure.
La consanguinité :
Dans ce cas, les individus se reproduisent entre membres
apparentés.
Dans ce cas, les deux individus ont une part de leurs gènes en commun. Au fil
des générations on tend vers une homozygotie de plus en plus complète. Mais
c'est beaucoup moins rapide que dans le cas de l'autogamie, qui est la forme
extrême de la consanguinité (les deux individus qui se reproduisent sont
alors vraiment très apparentés, puisqu'il s'agit d'eux-mêmes...).
L'homogamie :
Les individus se reproduisent avec des individus qui leur ressemblent
: rats noirs avec rats noirs, albinos avec albinos, etc. Deux cas de figure
:
- les gènes sont codominants : dans ce cas chacun se reproduit séparément :
AA avec AA, Aa avec Aa, aa avec aa. A chaque génération, le nombre de Aa
diminue de moitié, puisque ces accouplement produisent 1/4 de AA et 1/4 de
aa. Mais attention, seul le gène A est concerné...
- A est dominant par rapport à a. Dans ce cas, les AA et Aa se reproduisent
entre eux, et les aa se reproduisent entre eux. Occasionnellement, Aa x Aa va
produire quelques aa (qui rejoignent le second groupe). Dans ce cas de figure,
le groupe des A se purifie progressivement de l'allèle a, et le groupe aa
fonctionne en régime fermé.
Alors les individus ne sont pas nécessairement consanguins.
La consanguinité entraînant des similarités génotypiques, donc
phénotypiques, on comprend bien qu'elle est
tout de même nettement favorisée.
Ces trois régimes conduisent progressivement (à des vitesses différentes, malgré tout) à une homozygotie complète du génome, partiellement contrebalancée par les taux de mutations.
La panmixie :
Dans ce cas de figure, les reproductions se font de façon totalement
aléatoire, sans pression d'aucune sorte. Dans ce cas de figure, la structure
de la population reste plus ou moins stable, avec des génotypes fortement
hétérozygotes.
L'hétérogamie :
Il s'agit d'un cas très particulier : les unions se font entre
individus dissemblables uniquement. C'est l'inverse
exact de l'homogamie. Le cas le
plus simple et le plus évident est celui des chromosomes sexuels chez les
mammifères : les males se reproduisent uniquement avec des femelles, et
inversement. Dans ce cas, sur l'ensemble de la population, 1/4 des chromosomes
sexuels sont Y et 3/4 sont X (le male étant XY et la femelle XX), et ce
rapport est stable au fil des générations : selon que le père transmet (de
façon aléatoire) son chromosome X ou son chromosome Y, on obtient 1/2
femelle ou 1/2 male...
Ces schémas ont toutefois leurs limites... Chez l'homme, par exemple, il y a hétérogamie pour les chromosomes sexuels, panmixie pour les groupes sanguins, et une autogamie partielle (d'origine culturelle) pour la pigmentation de la peau. Il faut donc raisonner gène par gène. Mais la similarité pour un gène donné implique souvent la similarité pour d'autres gènes, qu'ils soient liés (présents sur le même chromosome) ou non, ne serait-ce que parce que des individus géographiquement très proches et d'aspect similaire ont des chances d'être partiellement apparentés. En outre la sélection va fausser le pur jeu du hasard, de même que la dérive génétique...
Une pression évolutive, c'est un phénomène externe qui va influer sur la structure génétique de la population et en modifier l'évolution théorique. Il en existe quatre : la mutation, qui fait apparaître de nouveaux gènes et allèles, la sélection, qui en favorise certains au détriment d'autres, la migration, qui introduit des génotypes nouveaux dans une population limitée, et enfin le hasard lui même, puisque l'effectif est limité et que les statistiques ne s'appliquent jamais à la lettre (une pièce lancée en l'air ne tombe pas à moitié sur pile et à moitié sur face :-) ).
Pour illustrer mon propos, nous allons considérer le cas d'un croisement entre Raoul, un male rex (Re/re) et Raoulette, une femelle lisse (re/re).
Le hasard et la dérive génétique :
|
Toutes les populations sont obligatoirement limitées. Dans le cas de
l'homme, la population maximale que l'on puisse envisager est de plus ou moins
6,5 milliards d'individus. A cette échelle, on peut considérer que la
population est à peu près infinie. Pour ceux qui ont du mal avec la notion de hasard, prenons le cas du sexe. Prenons notre exemple : Raoul rencontre Raoulette. En théorie, on
obtient 50% de Re/re (rex) et 50% de re/re (lisse). |
On voit que dans tous les cas de figure la fréquence de l'allèle Re a changé. Et elle change à chaque génération. C'est ce que l'on appelle la dérive génétique. Parce que l'on effectue un tirage au hasard dans un effectif fini (c'est à dire limité), il y a toujours un écart entre la théorie et la réalité. De
plus, plus un allèle est rare, plus il se raréfie (s'il n'est pas favorisé
par la sélection). Par conséquent, pour un gène donné, un des allèles va
toujours finir par s'imposer, à la longue. 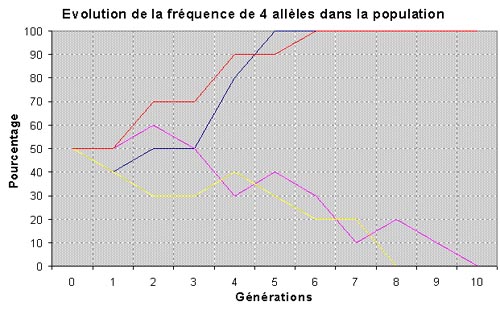
La conséquence de cette dérive génétique, liée au hasard, c'est que progressivement la population devient de plus en plus homozygote, parce que chaque gène subit ce phénomène. Plus la population est limitée, plus la dérive génétique est rapide. |
Goulot démographique et appauvrissement génétique :
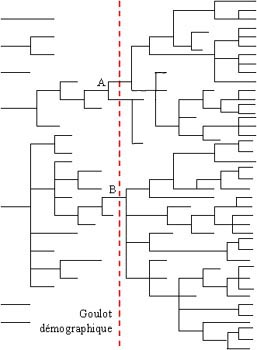 |
Plus l'effectif est limité, plus le nombre de gènes disponibles va
tendre à diminuer. De nombreux gènes disparaissent alors. Si par la suite la
situation s'améliore et que la population devient très nombreuse, cela ne
change guère le problème : les gènes sont hérités de nos ancêtres. Les
mutations étant peu fréquentes, même si la population passe de 10 à 10 000
elle n'aura pas 1000 fois plus de gènes, elle n'aura que les gènes transmis
par les 10 survivants d'origine. On le voit, si la dérive génétique semble indépendante de la consanguinité, en pratique elle y est généralement liée. Plus la population est limitée, plus deux individus de la génération N risquent d'être apparentés, parce qu'il n'y a pas un nombre illimité d'ancêtres possibles. De plus, tous les individus ne se reproduisent pas soit parce qu'ils ne vivent pas assez longtemps, soit parce qu'ils ne sont pas assez fertiles, ce qui diminue encore le nombre d'ancêtres possibles. Les gènes disparaissent donc à cause de la dérive génétique, mais aussi à cause de l'extinction d'une partie des branches généalogiques. Sur le schéma de gauche (simplifié: évidemment il faut un couple pour avoir des descendants, pas un seul individu :-) ), on part d'une population de 7 individus non apparentés, pour arriver à une population de 41 individus. Mais si l'on suit l'évolution de la population on se rend compte qu'à un moment donné la population a frôlé l'extinction, avec un minimum de 3 individus. Pire, tous les gènes présents dans la population finale proviennent de seulement 2 individus, A et B. Donc les 41 individus ont hérité d'un patrimoine génétique très limité : 2 ancêtres seulement, parmi les 7 de départ. Le patrimoine génétique global a donc diminué de 70% environ. Et encore, si on considère que l'intégralité du patrimoine génétique de ces individus s'est transmise à la population finale, ce qui est à peu près impossible... |
La sélection :
C'est justement la sélection qui provoque l'extinction de la plupart
des branches d'une famille, dans la nature comme en élevage. La sélection naturelle
tend à favoriser certains individus, certains génotypes. On parle
alors de valeur
sélective du génotype. Cette valeur comporte deux facteurs :
- la viabilité, qui est la probabilité de vivre suffisamment longtemps pour avoir une chance de se reproduire. Cette viabilité peut être liée à la prédation (les rats blancs vivent moins longtemps que les rats marron), au culling (tu es blanc, alors que je veux uniquement des marrons, hop, à la poubelle), à la santé (les rats épileptiques ont plus de mal à assurer leur survie que les rats sains, les rats immunodéficients sont plus souvent et plus gravement malades, les rats idiots s'empiffrent de mort-aux-rats...), etc. La viabilité, c'est la durée de vie de l'individu lui-même.
- la fécondité, qui est le nombre moyen de descendants par individu (le male le plus séduisant et le plus fort peut avoir facilement 2 000 descendants, le scrophuleux à roulettes aura bien du mal à en avoir une douzaine sur un malentendu...). La fécondité, c'est donc la durée de vie des gènes de l'individu dans la population. Plus un rat a de descendants, plus ses gènes se transmettent, moins ils risquent de s'éteindre rapidement (cf. paragraphes précédents)
La sélection naturelle comme la sélection humaine vont jouer sur ces deux
facteurs. Cependant, lorsque la première sélectionne les rats marrons, et la
seconde les rats dumbos violets, toutes deux favorisent un certain nombre de génotypes
au sein de la population. Tous les autres génotypes sont alors écartés, et
disparaissent plus ou moins rapidement. Les gènes récessifs peuvent
résister plusieurs générations, grâce aux porteurs hétérozygotes qui ne subissent
donc pas
les effets du gène mais peuvent le transmettre
à leurs descendants.
La sélection vient donc diminuer à la fois la population dans son ensemble
et la population reproductrice, c'est à dire le nombre d'ancêtres
disponibles pour les générations suivantes. La sélection (naturelle ou
humaine) augmente donc la consanguinité, dans une certaine mesure.
Cependant, on s'aperçoit que dans beaucoup de cas les génotypes
hétérozygotes sont les plus compétitifs, aussi bien pour leur viabilité
que pour leur fécondité. Ce dispositif permet de protéger les populations
contre une consanguinité excessive (cf. chapitres suivants)
|
Toutes
les mutations n'ont pas le même effet. Certaines ajoutent un morceau de code
génétique, d'autres en enlèvent, et enfin certaines le modifient simplement
sans modifier la quantité. De plus, un gène se compose de parties actives et de parties "muettes", inutiles ou à peu près inutiles. Certaines mutations n'ont donc aucun effet. Elles sont dites muettes, et passent totalement inaperçues tant qu'on ne pratique pas un séquençage génétique. Certaines ont un effet positif, et d'autres un effet négatif. Un gène, c'est la notice de fabrication d'une enzyme. Si la mutation améliore l'efficacité de l'enzyme entraînant un bénéfice pour le rat, la mutation est dite positive. Si au contraire elle diminue ou annule l'efficacité de l'enzyme, entraînant un handicap pour l'animal, même léger, elle est dite négative. Certaines modifications n'entraînent ni amélioration ni dégradation, elles sont donc neutres. |
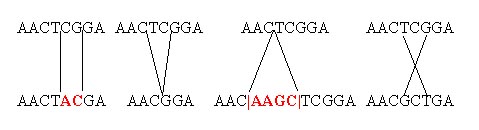 Différents types de mutation : erreur de copie, effacement, ajout, inversion... On pense qu'au moins une partie de l'ADN "inutile" est en fait formé des reliques de gènes rendus inefficaces par des mutations, sans mettre en péril la survie des individus mutants. |
Enfin, certaines modifications entraînent la mort du rat. Ces modification sont dites létales. La plupart du temps, les mutations létales entraînent la mort de l'embryon dès les premiers stades, et passent donc inaperçues. Mais parfois elles ne s'expriment qu'à partir d'un certain age. C'est le cas par exemple du gène Zitter, qui entraîne une dégénérescence du système nerveux, aux alentours de l'age du sevrage.
Il y a donc deux facteurs : le taux de mutation (nombre de mutations apparaissant à chaque génération), et la viabilité induite par ces mutations. Si la mutation est favorable, la sélection va entraîner sa diffusion au sein de la population. Si elle est neutre, elle sera soumise à la dérive génétique, et survivra ou disparaîtra. Si elle est négative ou létale, elle sera éliminée par la sélection.
Attention, la mutation est la seule source naturelle d'allèles nouveaux. Les mutations peuvent être induites par la radioactivité ou par certains produits, mais la très grande majorité apparaît au hasard. Certains pensent que des pressions évolutives peuvent augmenter la fréquence des mutations, mais à l'heure actuelle on est à peu près certains qu'elles ne jouent aucun rôle dans la "qualité" de la mutation elle-même.
La migration :
Chez le rat sauvage, et dans une certaine mesure chez le rat
domestique, la migration est un phénomène très important. C'est ce que l'on
appelle le "sang neuf". Il s'agit du transfert de gènes d'une
population vers une autre.
Les exemples les plus frappants sont observés chez des populations totalement
isolées, comme les reliques glaciaires. Ces animaux sont adaptés au froid. A
la fin de l'ère glaciaire, ils se sont réfugiés dans les montagnes au lieu
de migrer vers le nord. Mais
plus le climat devenait chaud, plus elles sont devenues isolées les unes des
autres.
On parle alors d'insularité : les populations sont isolées par la mer, par
le climat, par les barreaux d'une cage, etc. Il n'y a aucun échange
entre elles, aucun transfert de gènes. Chaque population va donc évoluer de
façon indépendante, sous l'influence des pressions évolutives citées
précédemment. Dérive génétique et consanguinité vont produire des
génotypes globalement très homozygotes, mais le plus souvent différents les
uns des autres à cause du hasard, d'un environnement légèrement
différent, ou de la sélection humaine.
Si on déplace un ou plusieurs individus du groupe A vers le groupe B, on
introduit dans le groupe B de nouveaux gènes, quelques uns réellement
nouveaux (apparus par mutation après la séparation des deux groupes), mais
pour la plupart disparus des générations auparavant. Plus les populations
sont génétiquement éloignées, plus l'effet est spectaculaire. On obtient
alors des "hybrides" très fortement hétérozygotes pendant les
quelques premières générations, puis de nouveau de plus en plus homozygotes
à nouveau. Mais les phénomènes de dérive génétique ont été bousculés,
et la sélection s'opère sur de nouveaux génotypes. L'ensemble
de la population B peut
donc être métamorphosée par l'ajout de quelques reproducteurs extérieurs
à peine.
La migration aura d'autant plus d'effet que les populations sont
éloignées, génétiquement et géographiquement, les deux étant plus ou
moins liés par une insularité partielle ou complète (même chez l'homme, il
y a plus de mariages au sein d'un même département qu'entre la France et la Papouasie...)
De même, plus les deux populations sont réduites, plus elles seront
consanguines, mais plus la migration d'un seul génotype aura d'effet. Sur des
grandes populations la migration a donc peu d'effet (tant qu'elle concerne peu
d'individus), mais de toute façon grâce à sa taille elle est déjà
relativement hétérozygote.
Enfin, on citera pour mémoire le génie génétique, qui permet d'introduire des gènes extérieurs non à une population mais à une espèce (c'est à dire la population ultime, dans un certain sens), par exemple un gène de haricot dans un saumon ou un gène de mouche dans un rat. Mais ces manipulations ne concernent pas le rat domestique et restent très rares, donc laissons les de coté...
![]() La consanguinité dans la nature :
La consanguinité dans la nature :
Le modèle théorique est la reproduction aléatoire au sein d'une
population illimitée. Dans ce cas de figure, les homozygotes sont très
rares. Mais en réalité dans la nature la reproduction est loin d'être
aléatoire. De nombreux dispositifs viennent fausser le jeu.
Il y a en premier lieu la sélection naturelle : les individus les moins bien
adaptés n'atteignent pas l'age de reproduction, ne rencontrent pas de
partenaire sexuel, ou sont supplantés par des concurrents mieux
adaptés. Par conséquent les rats sauvages ne se reproduisent pas au
hasard, et seule une petite proportion se reproduit.
Un autre problème est la taille de la population. Beaucoup d'espèces ont un
habitat discontinu, morcelé : l'espèce se compose de nombreuses populations
relativement (ou totalement) isolées les unes des autres.
De plus, chez la plupart des animaux sociaux, dont le rat, la structure familiale est fortement favorisée. Chaque groupe se compose de dominants qui se reproduisent, de leurs rejetons, et d'individus de statuts divers, mais pour la plupart apparentés au reste du groupe. Il est très rare que des individus non apparentés soient admis dans le groupe, et encore plus qu'ils parviennent à s'y reproduire. C'est pourquoi les rats sauvages sont souvent beaucoup plus consanguins qu'on pourrait l'imaginer de prime abord.
Évidemment, lorsque la consanguinité commence à entraîner des problèmes
de santé, la sélection naturelle vient faire son tri, et les rats étrangers
à la tribu ou les métisses deviennent "plus compétitifs"...
Dès cet instant, la consanguinité des individus retombe à zéro, ou presque
(rappel : lorsque deux très rats consanguins, mais non apparentés, se
reproduisent, leurs rejetons de la première génération sont apparentés,
mais pas consanguins : chaque rat a apporté des gènes différents. C'est
pourquoi deux rats homozygotes produisent des génotypes
hétérozygotes). Ce
taux de consanguinité va ensuite à nouveau augmenter progressivement. Comme
ces structures tribales familiales sont relativement petites (un seul à quelques
centaines d'individus), la migration joue un rôle très important dans
l'évolution de leur structure génétique.
Si on se place dans le cadre du rat domestique, on diminue l'impact de la sélection naturelle. Plus de prédation, un climat stable, des ressources alimentaires illimitées, une reproduction contrôlée, des soins médicaux... Il ne reste guère que des facteurs internes (stérilité, gène létal, etc.). Dans ce cadre, l'homme remplace la sélection naturelle. Attention toutefois, comme nous l'avons vu la sélection naturelle est loin d'être le seul facteur important... Il ne faut surtout pas minimiser le rôle du hasard et de la dérive génétique, surtout dans le cadre des ratteries qui disposent généralement (seulement) de 2 à 30 rats fertiles à un instant donné.
Reproduction aléatoireReproduction contrôlée
Lorsque l'homme intervient, il choisit les reproducteurs. La part du
hasard diminue considérablement. Par exemple, les agoutis se reproduisent uniquement avec les
agoutis et les noirs avec les noirs.
Lorsqu'un gène se retrouve sous une forme homozygote dans une famille, il est
dit fixé : en effet les descendants en hériteront forcément sous sa forme
homozygote, aussi longtemps qu'il n'y aura pas de brassage ou de mutation sur ce gène.
Par exemple, lorsque l'on reproduit uniquement des noirs, il ne peut plus apparaître
d'agoutis dans la famille, le gène ayant complètement disparu de la lignée.
Il est bien évident que de cette façon les gènes récessifs que l'homme souhaite sélectionner vont se multiplier très rapidement : en ne reproduisant que des rats homozygotes ou porteurs, il élimine les rats non porteurs du gène en question. Cependant, en favorisant des animaux présentant un caractère commun, donc souvent apparentés, l'homme augmente le taux de consanguinité.
Le choix des reproducteurs, donc la façon dont l'homme modifie la structure génétique de la population en diminuant la part de la reproduction aléatoire, c'est ça la sélection. Entendons-nous bien : la sélection ne crée rien. Par définition, elle sélectionne ce qui existe déjà, que ce soit visible ou caché. L'amélioration d'un caractère par la sélection a donc des limites, surtout quand il s'agit d'un gène simple. Et si l'on attend qu'apparaisse une mutation favorable, on risque d'attendre longtemps !
Par conséquent, il est vital de choisir attentivement ses reproducteurs. Surtout dans le cas d'une reproduction consanguine, l'erreur ne pardonne pas : la consanguinité va fixer de façon directe des gènes "positifs", ceux que l'on sélectionne, mais aussi des gènes négatifs, de façon indirecte, en augmentant la fréquence des homozygotes.
C'est pourquoi l'homme se substitue à la nature pour effectuer une sélection, sous diverses formes, la principale étant la sélection des reproducteurs. L'objectif est très simple : à partir de leurs caractéristiques et de celles de leurs ancêtres, les reproducteurs sont choisis de façon à fixer certains gènes "positifs" et éradiquer certains gènes "négatifs". Il s'agit donc d'une double sélection, positive et négative, la seconde éliminant les conséquences involontaires de la première.
N'oublions pas toutefois que la sélection ne s'opère généralement pas
sur le génotype directement, mais sur le phénotype, car on considère
généralement que le phénotype est en grande partie le reflet du génotype.
Mais tout n'est pas génétique et inné chez le rat domestique.
Au niveau comportemental, le rat apprend énormément, et ses tendances
innées peuvent donc être inhibées ou éliminées par l'acquis : un rat
nerveux peut devenir adorable s'il est bien éduqué, un rat paisible peut
devenir peureux à la suite d'un traumatisme...
Au niveau de la santé, quelques rares gènes ont un effet direct (épilepsie,
diabète, mégacolon, etc.). Mais la plupart ont un effet très indirect. Par
exemple, un défaut dans la structure du gène commandant les sites actifs des
récepteurs de certaines cellules immunitaires peut entraîner une certaine
fragilité face aux maladies. Des protéines pas assez
élastiques dans la paroi des artères peuvent
entraîner des problèmes cardiaques. Etc. Mais le fait de mourir jeune d'une
maladie ne signifie pas que l'on y était prédestiné. Une crise cardiaque
peut être liée à un terrain défavorable, mais peut aussi être liée à
une consommation abusive de mégots et de rillettes... ou à la simple
malchance ! Et que dire de problèmes dont la cause reste non
identifiée ? Une défaillance des reins ou du foie par exemple. problème
alimentaire, hygiène de vie, bactérie, séquelles d'un épisode infectieux, ou
génétique ?
Il importe donc de ne pas sauter trop vite aux conclusions. Et même en se basant sur les antécédents familiaux, la fiabilité du diagnostic "héréditaire ou non ?" reste limitée, à cause de l'effet du hasard que nous avons cité précédemment. Celui-ci ne joue pas seulement sur la répartition des gènes, mais sur tous les accidents de la vie, y compris les maladies, infectieuses ou non.
![]()
|
|
Les conséquences de la consanguinité |
Examinons un peu les conséquences de ce système de reproduction
dans quelques cas de figure...
La consanguinité a pour conséquence l'augmentation du nombre
de paires homozygotes dans le génotype de chaque génération successive,
nous l'avons vu. Cela signifie que les gènes récessifs, qui restent
généralement masqués dans une population idéale, ont beaucoup plus de
chances de s'exprimer :
|
Soit le gène albinos. Ce gène présente (entre autres) deux allèles : l'allèle sauvage dominant, C, sans effet. et l'allèle siamois, récessif, ch, qui produit des rats siamois sous sa forme homozygote. Les génotypes possibles sont :
Le rat A porte le gène siamois "ch". Il est hétérozygote C/ch.
On dit qu'il est porteur siamois. La ratte B, elle, ne porte pas le siamois. Elle est
homozygote C/C Ainsi de suite... On n'avait aucun rat siamois au départ, et on en a déjà un sur seize à la deuxième génération... Par contre la proportion de porteurs diminue progressivement (mais la quantité d'allèle ch reste constante : 8/32 soit 1/4, comme dans la génération F0) |
Cet exemple est bien anodin, car il permet simplement de faire surgir des
siamois là où il n'y en avait pas.
Mais cela se produit pour tous les gènes
récessifs que portent chacun des deux parents. Si l'un d'eux porte aussi un
gène récessif de diabète, et l'autre un gène récessif d'épilepsie, c'est
pareil, 1/8 des petits enfants seront diabétiques, 1/8 épileptiques (et donc
1/56 seront à la fois diabétiques et épileptiques). Plus grave encore, 50%
seront porteurs du diabète, et 50% seront porteurs de l'épilepsie. Donc un
quart des rats seront porteurs des deux à la fois !
C'est ce que l'on appelle la dépression consanguine. L'accumulation
de gènes récessifs finit par affaiblir progressivement l'organisme et entraîne l'apparition de toutes sortes de défauts : diminution de la taille,
baisse de fertilité,
faible résistance aux maladies, malformations, problèmes neurologiques
(comportement, perte d'acuité des sens, etc.)...
Certains sont liés à un gène en particulier, d'autres à des ensembles de
gènes. Pris indépendamment, chacun de ces gènes n'a pratiquement aucun
effet. Ils ont même parfois un effet positif. Mais ils sont en interaction
les uns avec les autres, et par "effet domino", la somme de ces
petites modifications peut produire des désordres majeurs. C'est ce que l'on
appelle des facteurs polygéniques. Ceux là sont très difficiles à
éradiquer. En effet, il est déjà difficile d'éradiquer avec certitude un
gène récessif simple. Donc en éradiquer plusieurs à la
fois c'est très
compliqué, il en reste toujours quelques uns, au moins masqués (le rat est
alors porteur d'une partie de ces gènes).
Attention toutefois, grâce à la sélection on peut contrebalancer en partie cet effet de dépression. En effet, en éliminant au fur et à mesure des programmes de reproduction tous les individus présentant des tares potentiellement héréditaires, on ralentit considérablement cet effet, même en maintenant des lignées très consanguines. De plus, la consanguinité favorise la sélection de traits positifs, comme la docilité, la taille, la résistance à certains types de tumeurs, etc.
Ainsi, beaucoup de souches de laboratoire sont des
souches fermées, c'est à dire consanguines. C'est par exemple le cas des Sprague-Dawley, un des principaux groupes de
souches, dont tous les représentants découlent d'un seul
et unique couple de rats originel (la souche Sprague-Dawley a été fermée en 1958, ce qui fait donc près de 50
ans de consanguinité). Et ce n'est pas la plus ancienne des
souches consanguines...
Il est donc parfaitement possible d'obtenir des souches consanguines viables,
sans défaut majeur. Mais attention, l'élevage des rats de laboratoire n'est
pas franchement "sentimental". Les rats de laboratoire ne sont
pas les chouchous à leur môman, ce sont des produits
industriels, généralement brevetés. Ceux qui présentent un défaut sont éliminés. La création des
souches de laboratoire
passe par le sacrifice de centaines de milliers de rats. Des milliers de
souches s'arrêtent dès les toutes premières générations, justement
lorsque les premiers soucis commencent à apparaître...
L'exemple des souches de labo est malgré tout
intéressant. Il existe deux types de souches de laboratoire :
|
On peut faire exactement le contraire de l'inbreeding. On appelle ça
l'outbreeding (ouah, dur...). Et ça signifie évidemment "reproduction
externe". Il s'agit de croiser deux rats qui ne sont pas
apparentés. L'objectif est de diminuer la consanguinité, en utilisant
l'effet d'hétérosis, ou vigueur hybride. L'outbreeding est
donc un régime de reproduction ouvert, équivalent domestique de la migration
évoquée plus haut.
On a vu que la consanguinité a tendance à augmenter
l'homozygotie sur tous les locus
( à la longue). Une fois parvenu à cette
homozygotie, on
obtient une lignée pure, c'est à dire une population où un seul allèle est
disponible pour chacun des gènes considérés.
|
Par exemple, une lignée pure d'albinos, c'est le type de lignée que l'on obtient en croisant deux albinos : chacun d'entre eux est génétiquement c/c, ils ne peuvent pas transmettre d'autres allèles pour le locus C : ni un allèle normal C, ni un allèle siamois ch ne peuvent apparaître, aussi longtemps qu'il n'y a pas de nouvelle mutation sur le locus C, qui viendrait en créer un nouveau. |
En sélectionnant sur un gène particulier par le biais de la
consanguinité, on purifie généralement le gène recherché (l'albinos, dans
l'exemple), mais aussi tous les autres gènes, de façon involontaire et (dans
une certaine mesure) incontrôlable. Cela entraîne généralement l'accumulation de tares
plus ou moins graves. Pour les éliminer, on a recours à la sélection. Mais
comme on l'a vu, une fois une qu'un gène est purifié par la dérive
génétique ou par la consanguinité, on ne peut plus obtenir d'autre allèle
que celui qui a "survécu".
Pour l'éliminer, on a alors recours à une deuxième lignée
pure. Cette lignée présente elle aussi les caractères souhaités, mais les
caractères indésirables ont subi une autre dérive génétique. Si on croise
les deux, on garde la lignée pure sur les allèles que les deux lignées ont
en commun, mais on brasse le reste du génome.
Admettons que l'on sélectionne une
lignée pure A et une lignée pure B. Les deux lignées sont
sélectionnées pour la docilité, l'albinisme, et le goût prononcé
pour la tarte aux pommes. Malheureusement, au fil des générations, la
lignée A a développé une petite taille, à cause d'une déficience
héréditaire en hormone de croissance (gène récessif). Tant que la lignée A reste fermée, consanguine, impossible d'obtenir
des rats de taille normale, car le gène "taille normale"
n'existe plus dans son patrimoine génétique : tonton Nestor était
le dernier à le trimballer, et il a sauté du 18è étage à l'age de
3 mois, emportant son gène avec lui... Si on croise la lignée A avec la lignée B, la
première génération sera obligatoirement docile, albinos, et aimera
la tarte aux pommes. En effet, où irait-elle chercher les gènes
"hargneux", "pas albinos" et "agreuh, aime pas la
tarte aux pommes, moi !" ? Ils n'existent ni dans la lignée A, ni
dans la lignée B... Cette première génération est donc albinos,
docile, aime la tarte aux pommes, mais ne présente aucune des tares
caractéristiques des lignées A et B. |
Évidemment, cette vigueur hybride est surtout manifeste à
la génération F1. Si on les croise entre eux ensuite, certaines des tares
vont de nouveau se manifester, puisqu'ils en sont tous porteurs. A la
génération F2, on obtient donc 1,6% de malchanceux qui ont hérité de
toutes les tares, 1,6% de chanceux qui ne portent plus aucune
tare, et une
majorité qui porte une ou plusieurs tares. En éliminant des programmes de
reproduction tous les individus qui présentent une ou plusieurs tares sous leur forme homozygote, on va
progressivement les éradiquer, même si ça prendra quelques générations.
Grâce à la sélection, à partir de deux lignées pures distinctes,
présentant chacune des défauts impossibles à éliminer à cause de la
consanguinité, on obtient à terme une nouvelle lignée pure qui ne
présente plus ces défauts. Évidemment, plus on utilise de lignées pures
distinctes, plus on élimine facilement les défauts.
Ca c'est la théorie. Malheureusement, elle se heurte à quelques problèmes...
- Il est généralement difficile de déterminer le degré de parenté avec
certitude. En gros, on considère généralement que deux rats qui n'ont
pas d'arrières grands-parents en commun ou apparentés (dans la limite des
informations disponibles) peuvent être considérés comme non apparentés. Ce
raisonnement est un peu faible, dans la mesure ou on
peut avoir facilement une vingtaine de générations à partir d'un seul et
unique couple de départ ( notamment en animalerie). Dans ce cas là, les deux rats sont en réalité très proches
parents, même s'il n'y a aucun ancêtre en
commun sur 10 générations... Mais bon, il faut bien une hypothèse de travail. Donc on choisit
des rats d'origine diverses (pas les mêmes animaleries, pas les mêmes
fournisseurs, etc.), et on espère qu'ils ne seront pas trop proches parents.
- Il suffit d'une seule génération pour annuler les effets
négatifs (ou positifs) de l'inbreeding, mais ces effets réapparaissent
ensuite et il faut les éliminer grâce à la sélection.
- La création d'une lignée pure implique que l'on possède de nombreux rats.
Deux lignées pures, encore plus. On risque donc d'être rapidement submergé
avant même d'être arriver à l'hybridation proprement dite...
- Sur le plan éthique, la création d'une lignée pure implique quand même
un certain nombre de générations de rats tarés.
- Certains gènes sont liés entre eux, et peuvent être ou devenir
indissociables. Dans ce cas l'hétérosis peut être impuissante à séparer
les traits recherchés des tares à éliminer.
Ce qu'il faut retenir principalement, c'est que la consanguinité ça n'est
pas héréditaire. Dès que l'on croise deux rats non apparentés, aussi
consanguins soient-ils, leur progéniture ne sera absolument pas consanguine.
Et ainsi de suite, aussi longtemps que l'on croise des rats qui ne sont pas
apparentés entre eux. S'il présente de gros atouts, un rat très consanguin présentant des défauts uniquement liés à cette
consanguinité peut donc être utilisé comme reproducteur sans le moindre
souci, dans le cadre de l'outbreeding.
A l'inverse toutefois, il ne faut pas oublier que si ses petits ne
présenteront pas de tares, ils en seront quand même porteurs. Il faudra donc
veiller par la suite à ne pas reproduire des rats apparentés issus de cette
lignée.
On l'a vu, la sélection ne peut pas créer de nouveaux gènes. Seule la
mutation peut le faire. Par conséquent, on ne peut sélectionner que les
gènes présents dans une population donnée. Le total de ces gènes
disponibles constitue ce que l'on appelle un pool génétique. Au fur et
à mesure de la purification des gènes, le pool génétique s'appauvrit. Les
mutations l'enrichissent très lentement. L'outbreeding, au contraire, vient
l'enrichir d'un seul coup.
La sélection va s'attacher à gérer ce pool génétique en choisissant
les individus qui vont assurer sa transmission aux générations suivantes. Le
pool génétique est donc en constante évolution, diminution ou
enrichissement.
![]()
|
|
Éléments d'éthique : Quels bons choix ? |
On l'a vu, pas de "vraie" lignée sans consanguinité. On parle plutôt alors de population, ou de descendants lorsqu'on se réfère à une filiation linéaire directe.
Cependant, il est important de garder le contrôle sur cette consanguinité. Pour prendre l'exemple des Prim'Holstein, il a été démontré que 1% de consanguinité en plus entraînait une diminution de la production de lait et une baisse de la fécondité, alors même que ce sont deux des principaux objectifs de la sélection chez cette race. On peut ainsi estimer un "taux de consanguinité optimum" au sein d'une population, permettant d'assurer un équilibre entre la dégénérescence par inbreeding et la sélection efficace des caractères recherchés.
Être opposé à la consanguinité en soi ne démontre que les
lacunes d'un éleveur. Il n'y a aucune raison d'être opposé à une
consanguinité raisonée, surtout chez une espèce stable, dont la population peut être
connue avec un minimum de précision. Cela est vrai chez le chien, le cheval
et la plupart des animaux anciennement domestiqués par l'homme, car dans ce
cas il est facile de se procurer des reproducteurs sains en se basant sur les
pedigrees, au lieu de "bâtards"
non identifiés.
Chez le rat on a une vaste majorité de "bâtards", issus de
reproduction aléatoire : les rats d'animalerie, entre autres.
C'est pour cela qu'il n'y a pas de race chez le rat domestique : même dans les pays anglo-saxons, les lignées ne sont pas suffisamment pures et anciennes pour prétendre à un tel titre.
Par contre, il ne faut pas tomber dans l'excès inverse et minimiser les
ravages de la consanguinité.
Chez le rat, on considère généralement que, en partant de rats non
apparentés (et on a vu qu'il est pratiquement impossible de le
déterminer), la dépression consanguine commence à se manifester de façon
significative au bout de 7 générations en moyenne. Cela ne signifie pas
qu'aucun rat de la 3è génération consanguine ne présentera de problème
lié à la consanguinité. Ce sont seulement des statistiques... En pratique,
on s'aperçoit que les problèmes apparaissent souvent dès la première
génération (cf. Paprika
x Massalé)
Si l'on se lance dans la sélection par lignées pures, on choisit consciemment de reproduire des rats qui présenteront des défauts plus ou moins graves, du fait de la dépression de consanguinité. On choisit donc aussi de faire adopter (ou alors de tuer) des rats qui auront des défauts plus ou moins graves : rachitisme, stérilité, fragilité immunitaire, malformations, etc. Ce choix doit être fait en connaissance de cause, et être clairement exprimé. En effet, il existe bien d'autres alternatives à ce modèle de sélection qui est à la fois efficace et moralement très discutable.
On pourra objecter que ces défauts n'apparaîtront que si ils sont déjà présents à l'origine, puisque "la sélection ne crée rien". C'est vrai. Alors, la consanguinité, ça ne pose pas de problème si les rats de départ sont sains ? Au contraire, il faudrait reproduire uniquement en consanguin, jusqu'à ce qu'on tombe sur un couple sain ? Malheureusement, tous les individus portent des gènes récessifs qui auront un impact négatif à une échelle ou une autre. Tôt ou tard, ces gènes récessifs finiront par être révélés par la consanguinité, car leur addition causera des déficiences qui ne seraient pas apparues en temps normal.
La plupart des ratteries opteront donc plutôt pour des modèles
intermédiaires, inspirés de
la sélection massale, moins efficace certes, mais beaucoup plus
"humaine". Car si l'éleveur a pour but d'améliorer les rats dans
les générations futures, il ne doit pas oublier qu'il est responsable du
bien-être des individus dont il a la charge. Choisir la souffrance d'un certain
nombre de générations dans l'espoir d'un bénéfice hypothétique, c'est une
politique douteuse.
En tous cas, de notre coté nous nous refusons à le faire. Il peut nous
arriver de faire appel à une consanguinité partielle dans certains cas, mais
nous avons su tirer les leçons du passé. Espérons que vous saurez tirer
les leçons de l'expérience des autres...
La consanguinité doit donc être maniée avec précaution, mais ne doit pas non plus être rejetée en bloc. A chacun de faire ses choix. On peut cependant émettre quelques suggestions : On considère souvent en France que l'on peut reproduire avec un risque raisonnablement faible deux rats dont le coefficient de parenté est inférieur à 1/16 (par exemple un seul des grands parents en commun, deux arrières grands parents en commun, une grand'mère et un arrière grand-père, etc.), à partir du moment où cela ne se reproduit pas plus d'une génération sur trois et si les familles ne présentent pas de défaut potentiellement héréditaire connu.
Ce modèle n'est toutefois pas le seul. Un des modèles courants dans l'élevage animal (tout type d'élevage confondu) est un système mixte, sur la base d'une consanguinité très élevée.
La méthode classique dans les élevages commerciaux consiste
à élever les animaux en consanguin (c'est à dire frère avec soeur ou père avec
fille) sur 5 à 10 générations, puis d'introduire un rat non apparenté, puis
recommencer.
Dans de nombreuses ratteries anglo-saxonnes, on alterne 2 générations de
consanguinité (frère-soeur ou père-fille), puis 3 générations d'élevage en
lignée et enfin une portée non consanguine. Ou alors exclusivement de l'élevage
en lignée.
![]()
|
|
| Bâtard | Issu de reproduction aléatoire, sans connaissance de ses ancêtres. Ce n'est pas un terme péjoratif, car les bâtards représentent un potentiel positif aussi bien que négatif. Mais ils représentent surtout une inconnue sur le plan de l'hérédité. |
| Chimère | La plupart des mutations sont éliminées de l'organisme. D'autres entraînent l'apparition de tumeurs. Lorsqu'une mutation se produit dans un organisme et se maintient ou se répand, on se retrouve avec un seul organisme composé de plusieurs génotypes différents. On appelle cela une chimère. Par exemple, certains rats noirs et blancs se retrouvent avec une zone de poils beiges, parce que la peau à cet endroit a subi une mutation. C'est aussi le cas de quelques rats vairons (odd-eyes). Les chimères ne sont pas héréditaires car elles se produisent dans des tissus somatiques (c'est à dire non liés à la reproduction) |
| Débile, taré |
Qui présente un défaut, quel qu'il soit, mineur ou majeur. Ca ne signifie pas nécessairement idiot ou subclaquant... Selon ce que l'éleveur considère comme un tare, ça peut être une paralysie totale ou simplement un marquage loupé. |
| Dominant, récessif, "codominant" |
Certains allèles s'expriment plus
intensément que d'autres. On dit alors qu'ils sont dominants
par rapport aux autres. Par exemple, l'allèle C (aucun effet) est
dominant par rapport à l'allèle c (albinos). A l'inverse, c est plus
faible que C. On dit que c est récessif par rapport à
C. Mais ces relations pas nécessairement aussi simples. Un allèle peut en masquer un autre, mais la combinaison de deux allèles peut aussi parfois produire un résultat intermédiaire. C'est le cas du gène hooded chez le rat, par exemple, pour lequel. D'autres s'expriment pratiquement à égalité (ce que l'on appelle couramment la "codominance"), comme l'albinos et le siamois qui produisent l'himalayen. |
| Gamète | Les gamètes sont des cellules sexuelles. Ce sont donc les gamètes qui assurent la transmission du patrimoine génétique à la génération suivante. Alors que le reste de l'individu est diploïde (les chromosomes vont par paires), les gamètes sont haploïdes (un seul chromosome pour chaque paire). La fécondation est l'addition des chromosomes des deux gamètes. L'œuf ainsi obtenu est à nouveau diploïde, et la moitié de ses gènes provenant de chacun de ses parents. |
| Gène, locus, allèle |
Un gène c'est un
"article" de l'information génétique. Un gène contient
toute l'information nécessaire à la fabrication d'une enzyme, par
exemple l'amylase salivaire. Ce gène peut être morcelé, réparti à plusieurs endroits sur un
même chromosome, voire sur des chromosomes différents. Ces endroits
portent le nom de locus.
Un locus peut avoir des "valeurs" différentes. Ces valeurs
sont appelées les allèles.
Chez les mammifères, les chromosomes vont par deux. Chaque chromosome présente la même structure, les mêmes locus, que son double. Par contre, les allèles ne sont pas nécessairement identiques. En effet, un des chromosomes vient du père, l'autre de la mère. Donc tous les gènes sont en double. Comme ils ne sont pas forcément identiques, on parle de chromosomes homologues et de gènes homologues |
| Génome, génotype, phénotype |
Le génome, c'est l'ensemble de
l'information génétique de l'individu, l'ensemble de tous ses
gènes. Cet ensemble définit le génotype de l'individu, sa carte d'identité génétique. Lorsque l'individu grandit, ce génotype s'exprime. Il s'y ajoute des influences externes : accidents, malnutrition, stress, etc. Tous ces facteurs produisent le phénotype, c'est à dire ce qui est visible, le résultat final. |
| Hérédité | Tout ce que les parents transmettent à leur descendance. Il s'agit des facteurs génétiques, bien sûr, mais aussi de l'éducation (par exemple la mère peut transmettre sa peur de l'homme aux petits sans qu'il y ait la moindre composante génétique) |
| Hétérozygote, homozygote |
Les gènes d'une même paire de
chromosomes sont dits homologues, mais peuvent avoir des valeurs
différentes. Si les allèles sont différents, alors on dit que le
rat est hétérozygote pour ce gène. Si les allèles sont identiques,
on dit qu'il est homozygote. Par exemple, c/c est homozygote albinos. C/c est hétérozygote (normal porteur albinos). c/ch est hétérozygote (albinos et siamois, c'est à dire himalayen) |
| Elevage en lignée (linebreeding) | Lorsque l'on cherche à créer une lignée, on prend un couple ou un individu de départ, que l'on considère comme idéal. Le but est de renforcer les caractéristiques de ce couple, éventuellement en en ajoutant d'autres. Pour cela on peut utiliser plusieurs méthodes. La première est de reproduire la père avec sa fille. La seconde, de reproduire un rat de la génération F2 avec sa cousine, de la génération F2. Dans tous les cas on favorise une augmentation de la consanguinité relative au couple ou à l'individu fondateur. Si l'on introduit des rats non apparentés, c'est uniquement pour inclure un ou plusieurs caractères dans la lignée, et dès la génération suivante on renforce à nouveau la part du génome du fondateur dans la lignée. |
| Mutation | Modification d'un gène, par le biais d'une erreur de transcription. Cette mutation crée alors un nouvel allèle, différent de l'allèle d'origine. Ce nouvel allèle peut être fonctionnel, ou au contraire être devenu inefficace. |
| Parthénogenèse | Reproduction sans fécondation : les
femelles peuvent produire des oeufs fertiles sans intervention d'un
male. Attention, les descendants ne sont pas des clones, il y a quand
même un certain brassage. Mais seuls les gènes d'un seul individu
entrent en jeu, ceux de la mère. La parthénogenèse est un mode de reproduction peu répandu. On la rencontre chez divers insectes, comme les pucerons ou les phasmes. |
| Population | Ensemble d'individus, quelle qu'en soit l'échelle : population humaine, française, bretonne, quimpéroise, famille Cadiou, etc. Dans une ratterie, on a une population totale, une population de reproducteurs potentiels, une population de males, etc. |
Le contenu de ce site : images et textes, ne sont pas libre d'utilisation, vous devez toujours me demander avant de les utiliser